
Webbia 56(2): 229-240. 2001
Paeonia morisii sp. nov. (Paeoniaceae), a new species from Sardinia
G. Cesca, L. Bernardo e N.G. Passalacqua
Orto Botanico dell'Universita della Calabria, Arcavacata di Rende, 87036 Rcnde (Cosenza)
Ricevuto il 9 Febbraio 2001 Accettata il 20 Giugno 2001
Paeonia morisii sp. nov., a new species from Sardinia. — A new species oi Paeonia from Sardinia is described. It differs morphologically, ecologically and karyologically from P. mascula subsp. russoi from Sicily, with which it was so far confused, as well as from other peonies of the P. mascula group.
Key words. Corse, Paeonia, Paeoniaceae, Sardinia, taxonomy.
According to actual Floras indications, Sardinian populations of Paeonia are referable to Paeonia mascula Mill, subsp. russoi (Biv.) Cullen & Heywood, which is a subspecies occurring in Sicily and Greece. The record of P. coriacea Boiss. from Sardinia seems to be due to a past error in picking up and arranging herbarium materials (Cullen & Heywood, 1964).
Yet since the first look, all the populations we studied appeared as homogenic among themselves and different from those observed in Sicily. The diversity founded upon either morphological (size, consistence and colour of leaves and stalk; number and shape of leaflets; shape and hairiness of fruits) or ecological characters.
A comparative morphological and karyological study revealed sufficient to separate Sicilian populations from Sardinian ones, and to propose the taxonomic acknowledgement as new species for the latter.
Paeonia morisii Cesca, Bernardo & Passalacqua, sp. nov.
Diagnosis — Planta perennis, erbacea, simplex, 25 - 60 cm alia. Caulis erectus, teres, subangulatus, glaber, paullo vel omnino purpurascens. Folia 4 - 7, alterna, inferiors 18 - 30 x 16 - 28 cm, biternata, segmentis (8-) 9 (-10) (raro 7 - 20), summa gradatim reducta dein segmentis 3 (-1); foliola coriacea, elliptico-ovata, 3 -12 x 2 - 9 cm, apiculata, basi inequilatera, breviter angustata, supra lucida, glabra, obscure viridia, subtus glauca, sparse pilosa usque lanosa, pilis longis crispatis- petiolis supra vel ambo latere purpurascentibus, superne saepe reductis. Flos plerumque unicus (raro 2 - 3), terminalis; calix zygomorphus, in fructu reflexus, sepalis 3, viridibus saepe purpurascentibus, extra pubescentibus; petala (5-) 6 - 8 (-9), obovata, purpurea, rosea vel albido- variegata (raro). Folliculi 3 - 5 (raro 2 - 7), irregulares, e medio ad apicem angustati, in stigmatem stylo tenui suffultum desinentes, suberecti, maturitate leviter reclinati, virides vel purpurascentes, tomento lutescente tecti.
A P. mascula (L.) Mill, et P. russoi Biv. pilis crispatis, foliis coriaceis et follicolis maturis minoribus superne angustioribus differt; a P. coriacea Boiss. et P. cambessedeii (Willk.) Willk. folliculis glabris differt.
Typus — "Monte D'Iscudu (Gennargentu, Nuoro, Sardegna), gariga a Genista Corsica e Rosa serafini ,1300 m, su calcare, 22. VI. 1996, L. Bernardo et N.G. Passalacqua s. n.", holotypus (CLU); Isotypi in Fi, PAL, SS
Etymologia — Species Magistro G. G. Moris. (1796 - 1869), florae sardae perspicaci investigatori, grata memoria dicata.
Synonima — P. corallina var. pubescens Moris (Moris, 1837); P corallina var. pubescens f. hypoleuca Briq. (BRIQUET, 1910).
Description — Plant herbaceous, perennial, monocaulis, 25-60 cm (rarely less) high, generally bearing a solitary flower. Root tapering, fleshy, simple or branched (1 or 2 branches). Stem erect, cylindrical or sub-angular, glabrous or sub-glabrous, with few sheaths more or less expanded at the base; the color may be varying from green and becoming purplish only in the lower parts (rarely completely green) to totally purplish. Leaves 4-7, alternate, cordate, coriaceous; lower leaves 18-30x 16-28 cm, biternate, with 9 leaflets (rarely 7-20); upper leaves progressively reduce till 3(1); leaflets vary from elliptic to ovate, 3-12 x 2-9 cm long, acuminate, usually asymmetrical and decurrent at the base; upper face deep green, glabrous and bright; lower face blue-green, more or less covered with long rippled hair. Leaf-stalk 1-10 cm, sub-glabrous, more or less purplish on the upper face, sometimes totally purplish; leaf-stalk of leaflets usually present in lower leaves, sometimes reducing (even lacking) in the middle ones, very reduced or absent in the upper leaves. Flower generally 1 (rarely 2 or 3), terminal, with 1 (3) bract similar to the leaflets of the upper leave. Calyx zygomorphic, covering the petals at the blossom, reflexed at the fructification; sepals 3, green (sometimes purplish) with outer face more or less tomentouse. Corolla spirocyclic with (5)6-8(9) ovate petals, pinkish mauve or white-striped, falling precociously. Stamens with yellow anthers and white to red filaments. Carpels (2-)3-5, tomentouse by white to whitish thick and short hair; stigma red, flattened and recurving. Follicles 3-5, irregularly shaped, tapering in the upper half and ending with the stigma carried by a thin style, sub-erect, curved backward when mature, from green to red and covered with yellowish tomentum.
Iconographia — Moris (1: 64, t. 4. 1837); Camarda. et al. (1992, sub P. mascula (L.) Mill, subsp. russoi (Biv.) Cullen & Heywood).
Biological form — Rhizomatous geophyte. flowering — March- April.
Chromosome number — 2n=10; these plants are characterized by a diploid chromosomical number (cfr. figs. 1, 2, 3 ; tab. 1).
The study of karyotype showed a prevalence of m (metacentric) chromosomes, with sm (sub metacentric) and st (sub telocentric) chromosomes too. Satellites have been observed on the short arm of sm and st; on the contrary, there is no evidence of satellites on metacentric chromosomes. We did not notice M (perfectly median centromere) chromosomes and t (telocentric) chromosomes: in fact, we consider the presence of an M chromosome in the plate of fig. 2 not to be significative.

Figs. 1-2 — Metaphase plates from root rip (x 1800 ca.).
table 1 — Mean values of measurement (micro-m) based on eight plates. BL = long arm; BC = short arm; L% = percentage length; i - centromeric index x=5=3m+1sm°+1st°

|
|
|
I
|
|
II
|
III
|
|
|
1-2
|
3-4
|
5-6
|
7-8
|
9-10
|
|
BL
|
10.8
|
8.95
|
8.36
|
9.6
|
10.7
|
|
BC
|
9.15
|
8.06
|
6.69
|
4.64+0.8s
|
2.14+0.8s
|
|
L%
|
24.73
|
21.11
|
18.66
|
18.55
|
16.91
|
|
BL/BC
|
1.18
|
1.11
|
1.25
|
2.07
|
5
|
|
i
|
45.86
|
47.38
|
44.45
|
32.58
|
16.66
|
The Levan's formula we elaborate on available plates is 2tt-10=6m+2sm°+2st°. The values of aploid idiogram are reported in tab. 1 and fig. 4.
Fig. 4 Haploid idiogram.

Ecology — Paeonia morisii grows in different kind of woods (more or less clear) but even in meadowlands, in stony grounds and among bushes above 400 m. It can be found on every type of geological substrate (either on calcareous or siliceous rocks, sedimentary and magmatic, metamorphic or not), and it grows also on a very poor soil, even if it prefers deep, rich and wet soils.
Distribution — Our description is based on Sardinian populations, the richest of the species; the observation of herbarium specimens shows that the species is present in Corse too, where either Paeonia mascula Mill, subsp. russoi (Biv.) Cullen & Heywood or P. corallina Retz. var. pubescens Moris. have been recorded.
Specimina visa selecta
Italy— Sardegna: Gennargentu -----1984 (VER) — Sardinia provincia di Sassari: ---- (FI, PAD, PT, RO) — In silvis montanis ex Sardinia, 1826, Moris (BOLO) — Montagna di Atla dei Sarda, 13-14. VI.1898, Martelli (PI) — Insula Sardinia, Reg centr. or. in Monte Oliena,., 12.V.1884, C. Forsyth (PI) — In pratis montanis editis prope Pulam Sardinia,, IV, U I. Maller (FI) — Gennargentu a Girgini, 2.VI.1896, Martelli (FI) — In insula Tavolara rupestribus calcareis rocce della Madonna, s. Coll.,25.V.1885 (FI) — Italia, Sardaigne, prov. Nuoro, Gennargentu, ---- (FI) — Arrondissement de Tempio. Monte Limbardo, maquis detouverts sur granit, 16.V et 14 VII.1882, E. Reverchon (FI) — Mti d'Aritzo, s. coll., VII. 1859 (GAG) — Monti Urtinu (CAG) — Gennargentu Arcu D'Iscova, 21.IV. 1948, Martinoli (GAG) — Cala Gonone, 22.V.1975, G. Milia et I. Camarda (CAG) — Desulo, Su Au, IV.1983, Goi A. (CAG), Saramini (Villagrande Strisalis), 18.V. 1983, Floreddu (CAG) — Mte Linas, 1984, Chiappini et Angiolino (CAG) — Acquafrida,, 20.1V.1986, M. G. Biagini (CAG) — Mte Tonneri-Semi, VI. 1987, M. Micro (CAG) — Punt a Sebbera, Domus De Maria (Cagliari), IV.1988, A R. Cottu (CAG) — Talasaggia, Tonara (Nuoro), V.1990, Fogu C. (CAG) — Bosco lungo la strada nel vallone Rio Arpfu (Fonni, Nuoro, Sardegna), Bosco di leccio c roverella, 1100 m, 22.VI.1996, L Bernardo et N.G Passalacqua (CLU) — Serra Edele-Bosco di Ghivine (Cala Gonone, Nuoro, Sardegna), lecceta aperta, in radure, 540 m, su calcare, 23.VI.1996, L. Bernardo et N.G. Passalacqua (CLUI — Monte Bruncu Spina (Gennargentu, Nuoro, Sardegna), piccolo bosco ripario ad Alnus glutinasa, 1600 m, su calcare, 22.VI.1996, L. Bernardo et N.G. Passalacqua (CLU) — Monte Albo (Lula, Nuoro, Sardegna), bosco di leccio, 1000m ca. su calcare, 21.VI.1996, L. Bernardo et N.G. Passalacqua (CLU).
France — Corsica: Serra di Scopamene, prés Sarthène (Corse), 4.VI. 1879, E. Reverchon (FI), — Pointe de Golfi-doni et Monte Fornello, versante est, maquis, silice, Briquet, 400-575, 27.IV.1907(GE).
Systematic considerations
Peonies are characterized by a particularly plastic phenotype, thus it is not simple to identify single morphological characters for distinguishing clearly two entities. It is more useful to refer to the variability range of a character and to the way the different characters join together in individuals, than to refer to a single character. Sardinian populations are a lot less variable than those of Sicily and Peninsular Italy, which can be assigned to P. mascula s. l. This element takes part in our taxonomic valuation together with karyological data.
In the next part of the work, we put in evidence the morphological, karyological and ecological characters we consider significant for the differentiation from P. mascula Mill, subsp. mascula and subsp. russoi (Biv.) Cullen & Heywood, which actually are the reference taxa for Sardinian population, since they are the most alike.
Moreover, we want to specify that we were not able to find the typus of P. mascula subsp. russoi (Biv.) Cullen & Heywood, so that we had to base the comparison with this taxon on bibliographical data and on observations of Sicilian material (locus classicus material included).
In order to obtain an appropriate comparison, we examined also the Italian herbarium specimens of Sicilian P. mascula subsp. russoi (in BO, CLU, FI, NA, PAL, PI, RO, VE) and of peninsular Italian P. mascula subsp. mascula (in BO, CLU, FI, PI, RO).
Morphology
Habitus — Sardinian individuals are usually less high and voluminous than Paeonia mascula s. l. They rarely branch out at the base, and leaves do not come off much from the stalk (fig. 5).

Fig. 5 — P. morisii in flower.


Fig. 6 — P. morisii. lower surface of leaflet.


Fig. 7 — Fruits of P. morisii.
Stem — The new species is characterized by an evidently purplish and coriaceous stem (figs. 6, 7), while the consistence is less tough in both the subspecies of P. mascula; moreover, the red stripes are absent or present mostly at the base.
Leaves — The number of leaflets is always very unsteady in both the subspecies of P. mascula: there are from 3 to 27 segments in middle and lower leaves, with an average of 12-15. Sardinian Paeonia shows a variability from 7 to 20 in middle and lower leaves and from 1 to 4 in the upper, but the number is almost always 9 in middle and lower leaves, and 3 in the upper. As regards the shape and the size of segments, the new species is characterized by a less marked variability than that of the two P. mascula subspecies, and the segments are usually smaller. The leaves are thicker in the new species, and the upper surface is brighter.
The hairiness is one of the most significant characters of Sardinian populations; in fact, it distinguishes for quantity and quality from that of the two subspecies of P. mascula. P. morisii has ever long and crinkled hairs, which are twisted and irregularly bent on different levels; it is abundantly placed on lower surface of foliar leaflets, even if some individuals can occasionally show a smaller or bigger reduction of hairiness. On the other hand, P. mascula Mill, subsp. mascula is always glabrous or subglabrous, and the subsp. russoi (Biv.) Cullen & Heywood has leaves, which can vary from subglabrous to tomentouse, with either long or short hair, but always right and quite regularly arranged (figs. 8-11).
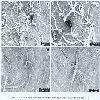
Figs. 8-11 —SEM view of hairs oflower surface of leaflets: P. morisii. (figs. 8-9) and P. mascula subsp. russoi (figs. 10-11).
Fruit — In P. mascula s. l. follicles are never purplish, more regular and bigger than in P. morisii; moreover, they show a rounded apex and curve more backward.
It is easy to discriminate the new species from P. coriacea Boiss. and P. cambessedesii (Willk.) Willk., because in the last two follicles are glabrous, whereas in P. morisii they are densely covered by short hairs (cf. fig. 7).
Karyology
Sardinian plants are characterized by a 2n=10 chromosomical number; this distinguishes them well from material of P. mascula s.l. (2n=20) analysed until now, both from Sicily (Raimondo et al., 1983) and peninsular Italy (Bernardo et al., 1995; L. Peruzzi, pers. com.). Many literary dates are difficultly interpretable, since they are apparently contradictory: Barber (1941) and Stern et al. (1944) indicated a 2n=20 for Sicilian, Sardinian and Corse P. russoi, without specifying which material was examined; the same for Cullen & Heywood (1964), which indicated a 2n=20 for P. russoi of Western Mediterranean Islands; Tzanoudakis (1983) indicated a 2n=10 for Greek P. russoi and 2n=20 for Greek P. mascula; at last, Sopova (1971) in Macedonia and Koeva & Sarcova (1993) in Bulgaria indicated 2n=10 for P. mascula.
Ecology
Paeonia morisii shows a wide ecological plasticity, from woody habitat to occasional open ground, unlike both the subspecies of P. mascula that occurs nearly exclusively in woods.
Acknowledgement
We wish to thank Directors and Keepers of mentioned Herbaria who have kindly provided us with the exiccata of studied species; moreover, we are grateful to Prof. Ignazio Camarda and Dr. Vincenzo Satta for the informations and the useful suggestions they have given us; finally our thanks to Jessica Barbieri for the translation help.
BIBLIOGRAPHY
Anderson G., 1818. — A monograph of the genus Paeonia. Transactions of Linnean Society of London 12: 248-290.
Barber H. N., 1941. — Evolution of the genus Paeonia. Nature 148:221.
Bernardo L., Bruno F., Cesca G. & Passalacqua N.G., 1995 — Specie critiche delta flora calabra: problemi sistematici e nuove segnalazioni. Boll. Soc. Sarda Sci. Nat. JO: 435-445.
Briquet J., 1910. — Prodrome de la Flore Corse. 1: 580-581, 651, Geneva & Basel.
Camarda I, Corrias B., Diana S. & Valsecchi F., 1992. — Piante di Sardegna. Ed. Chiarella, Sassari.
Cullen J. & Heywood V. Notes on the European Species of Paeonia Feddes Repertorium 69: 32-35.
Huth E., 1891. — Monographie der Gattung Paeonia. Botanische Jahrbücher für Systematie 14: 258-276.
Koeva J. & Sarcova S., 1997. — Karyological study of three species of Paeonia (Paeoniaceae) in Bulgaria. Bocconea J: 553-556.
Moris G. G., 1837. — Flora Sardoa 1:64 t.4. Torino.
Raimondo F. M., Rossitto M & Ottonello D., 1983. — Numeri cromosomici per la Flora Italiana: 984. Inform. Bot. It. 15:180.
Sopova M., 1971. — The cytological study of two Paeonia species from Macedonia Fragm. Balk Mus Macedon.Sci. Nat. 8(16): 137-142.
Stearn W. T. & Davis P. H., 1984. — Peonies of Greece. The Goulandris Natural History Museum, Kifissia.
Stern F. C., 1944. — Geographical distribution of the genus Paeonia. Proc. Lin. Soc. London 13:16-80.
Stern F. C., 1946. — A study of the genus Paeonia. London.
Tzanoudakis D., 1983. — Karyotypes of four wild Paeonia species from Greece Nord. J. Bot 3:307- 318.